01 Dec 2016
生物学者のDoping博士は、生きた細胞、分子、およびヌクレオソームフィブリルを凍結する技術を固定することについて語っています。
私たちはポストゲノム時代に生きています。 誰もがゲノムの配列を決定し、自分の細胞内にある遺伝子に関する完全な情報を得ることができます。 1つの細胞は46のヒト染色体を含み、約2メートルのDNA分子(各染色体はDNA分子を含む)の全長を含む。 実際、それは非常に長くて薄い分子です。 各細胞核に2mのDNAが小規模にパッケージされています。 約10ミクロンの核の大きさ。 細胞内でのDNAの圧縮は、約10,000倍であると考えられています。つまり、分子がコンパクト化された素晴らしい方法です。 逆説は、ゲノムが特に難しいとは読まなければ、この遺伝子がどのようにして細胞内でどのようにしてコンパクト化されているのかまだ分かりません。 確かに1世紀以上にわたって研究されてきたこの問題は、解決されていない。 細胞核の構成や有糸分裂染色体におけるDNAコンパクションの過程を根本的に変えなければならないとき、定期的に物語が起こります。
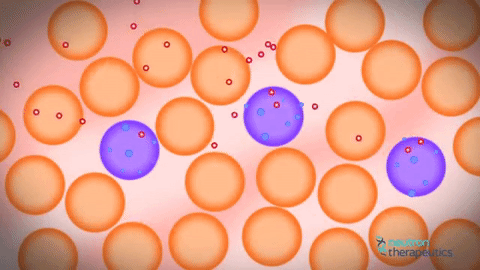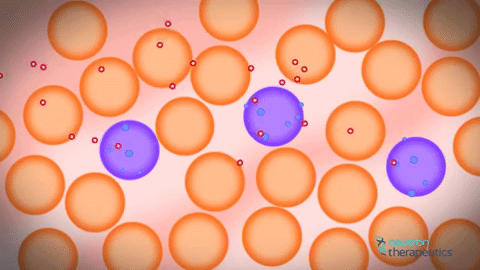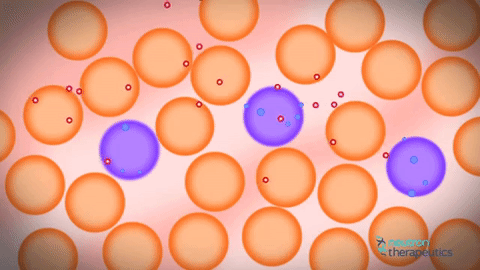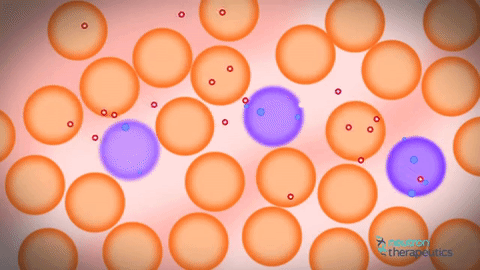
おそらく一貫してますます厚い原線維の形成と関連しているDNA分子の圧縮の基本原理。 分子は細かい原線維に最初にコンパクト化され、その後、この細いフィブリルはより厚く、さらに厚くなり、ある時点で有糸分裂染色体に変わる。 最近まで、これらのレベルは多かれ少なかれ説明されていたようでした。 DNA分子の第1の圧縮レベルは、いわゆるヒストンタンパク質との相互作用に関連する。 ヒストンタンパク質には2つの主要なグループがある。 コアヒストン - このヒストンH2A、H2B、H3およびH4は、これらのヒストンのそれぞれ2分子がホッケーパックのような形状のタンパク質構造を形成する。 そしてホッケーパックはDNA分子を傷つけました。 この薄い薄い2ナノメートルのDNA分子は、この洗濯機の回りに1,7回回転するとリンカー部位に行き、次の洗浄機などになります。 この構造は、電子顕微鏡で見ることができ、「紐上のビーズ」と呼ばれ、約10ナノメートルのフィブリルの厚さであった。 それは非常に長い時間、DNA分子の圧縮の最初のレベルと信じられていました。
ビタミンB12シアノコバラミン注射は - DNA合成に必須です。
興味深いことに、研究の初めに、電子顕微鏡(DNAコンパクションを研究することが不可能ではない)があったときに、これが主で唯一の原線維であり、何らかの形で染色体が形成されたと推定された。 それは研究の方法と関連していた。 電子顕微鏡は生きている細胞を見ることができません。 物体は非常に薄くなければならないので、電子は厚い物体を通って飛ぶことができない。 したがって、死んだ標本のみを見ることができます。 どのように死者のサンプルを作るかは、実験技術の状態に依存します。 当然のことながら、すべての成分が適所にあり、生きた細胞内でそれに特有の特徴を最も多く保持している必要があります。 この目的のために、ケージに注がれる様々な液体が細胞を殺し、理想的には、すべての成分が生きた細胞内の固定された状態である - この固定手順。
電子顕微鏡で使用された最初の良好なクランプの1つは、四酸化オスミウムであった。 それは脂質をよく捕捉し、核酸、タンパク質を非常に弱く結合するため、細胞内に10ナノメートルフィブリルが見られます。 アルデヒドクランプがより成功し、すでにタンパク質と相互作用することができました。 彼らは実際に、そのようなネットワーク内のすべての内部細胞空間を縫合し、互いに隣り合っているタンパク質を連結した。 そして、これらのロックは成功以上のものです。 クリップにアルデヒドを使用すると、染色体の核に10ナノメートルのフィブリルが見つからないことが判明し、クロマチン基本フィブリルと呼ばれるより厚いフィブリルが見つかった。 その厚さは約25〜30ナノメートルである。 そして数十年間、これはフィブリルがDNAコンパクションの基本ビルディングブロックであると信じられていました。
アルデヒドなどを使用して固定するときは、化学的固定であり、細胞はすべてそれを理解していますが、さまざまな変化があり、誤った情報の量は十分です。 あるタンパク質の固定時に、その位置が変化したり、抽出されて細胞が残ったりすることもあります。 今や、生体内観察の技術があり、生きた細胞のそれと、そして固定後のそれと比較することができます。 固定すると、多くのアーティファクトが発生します。 したがって、常に電子顕微鏡法を含むより多くのネイティブなクロマチン構造を探索する方法を見出したいと考えています。 しかし、電子顕微鏡法は?
細胞全体が正確にインビボであることができないことを見る。 ケージは、非常に薄いスライス(30,50,70,100ナノメートル)でカットする必要がありますが、厚い、または電子はストライクではありません。 しかし、電子顕微鏡でセルで多かれ少なかれネイティブに見ることを可能にした技術が開発されている。 そしてそれはすでに化学的な固定剤や物理的な固定、すなわち凍結につながっていません。 理論的には、細胞は水泡であり、そこには異なる分子が浮遊しています。それを凍らせると、おそらく大丈夫でしょう。 この問題はよく知られています。凍結中に極端に破壊された氷結晶が形成されます(生体分子は一般に非常に柔らかい)。 しかし、ガラス化した状態で水を凍結して移すことはできません。 非常に高速で凍結すると、氷の結晶は形成する時間がなく、実際には液体の水はただ安定しています。 分子は速い速度でストップを拡散し、構造全体が安定し、実際には硬いが、水には結晶がない。 問題はそのような状態で水を動かすことでした。
このような状態で翻訳する最も簡単な方法 - 非常に小さな液滴をとり、急速に冷却すること。 これが行われれば、それは液滴中のその固有の構造を維持することである。 ビトリファイド状態のサンプルを見ると、特定のオルガネラを実際にどのように見えるか、DNA分子の構造がコンパクト化されていることがわかるはずです。 それは非常に難しく、技術は非常に長い時間を進化させました。 これは、高分子複合体を含むサンプル溶液が驚異的なスピードで液状タンに還元されるため、様々なタンパク質分子、複合体、ウイルス、リボソームを探索するのに非常に積極的に使用される最も簡単な方法です。 温度は低く、非常に迅速に凍結されます。
しかし、細胞の場合、少なくとも動物細胞では、この方法の作業ははるかに悪いです。 それでも、彼らはそのようなアプローチのために素晴らしいです。 しかし、この技術が開発され、大細胞、例えばヒト細胞を凍結させる方法が提案されている。 1つのアプローチは以下の通りである:サンプルは同時に非常に速く冷却され、高圧ゾーンに入る。 凍結中の高圧(既知のように、凍結水がわずかに膨張し、氷密度が水の密度よりも小さい場合)は容積を増加させず、その結果水は結晶化せず、ガラス化状態で貯蔵される。 言い換えれば、それは非常にシンプルですが、実際には複雑な楽器であり、その上で頑張って良い結果を得ています。それはアートの芸術です。
これが凍っていると、サンプルがビトリファイド状態になります。つまり、物理的なロックがあります。 大きなサンプルはできるだけ凍結されていますが、組織の微視的ビット、組織さえあれば、個々の細胞はもちろん可能です。 それで、氷は冷凍して細かく細かく細かく刻むことができます。 当然ながら、それはまた、凍結切削を生産することができる特別な装置でもある。 さらに凍結切片を電子顕微鏡に移すことができます(モデルは常に液体窒素で冷却しなければなりません)。その後、電子顕微鏡で事実上元の状態で見ることができます。 もちろん、この国家がどのようにネイティブに議論しているかについて。 理論的には、もし我々が凍った細胞であって、氷結晶の形成なしにそれを非常にうまく解凍すれば、それは復活して生き続け、異なる物質を合成するなどであろう。 しかし、残念ながら、それは動作しませんが、それがネイティブ状態であることが期待されます。 このようなカットを見て、間質核を研究したところ、クロマチンが様々にコンパクト化され、様々なコンパクト化されたクロマチンが強くコンパクト化され、活性なRNA合成である拡散性クロマチンとそれ以下のコンパクションレベルがあり、驚くべきことに、他のサンプルのクロマチン原線維が見つからないということはありませんでした。 それは簡単ではありませんでした。 ヌクレオソームははっきりと見え、30ナノメートルのクロマチン原線維はない。
状況は、染色体の圧縮を研究する初期の段階とは正反対になっています。 その後、最初に、ヌクレオソーム10ナノメートルフィブリル、それ以外はないと考えました。 それから、それは事実ではないことが判明し、人々は議論し、議論し、記事を書きました。 彼らは結論として、おそらくヌクレオソームフィブリルのみであるが、基本的なクロマチン原線維が存在するという結論に達した。 そして、ここでは、その基本的なクロマチン小繊維に戻ることが必要であることが判明しました - それは、アーティファクトではなく、細かいフィブリルヌクレオソームがあります。
私の人生すべてを信じている人々にとっては、2つのレベルの圧迫があり、そのうちの1つがそうではないことが判明したのは、とてもイライラしています。 さらに、個体がインビボヌクレオソームをモニターすることが可能になったことが判明した。 不安定な核内のヌクレオソームは、少し動いて拡散することが判明した。 それからアイデアがありました、なぜこの基本的なクロマチンフィブリルを見ないのですか? 彼らがお互いの近くに位置していた場合、基本的なクロマチンフィブリル。 ヌクレオソームは動いており、どのようにして相互に浸透することができるだろう。 また、ヒストンタンパク質の相互作用はヌクレオソームだけでなく、個々のフィブリル間でも間接的に起こることが示唆されているにもかかわらず、クロマチン基本フィブリルの欠如が示唆される。 原則として、これはよく知られている事実です。 さらに、コンピュータクロマチンの圧縮をシミュレートしようとするとき、それは非常に積極的に方向を開発しています。 これはかなり複雑なプロセスで、スーパーコンピュータを使用する必要があります。 コンピュータで計算することはまだできません。そのため、DNA分子をコンパクト化する必要があります。
一方では、はい、古いの拒絶、他の新しい機会。 これは、フィブリルの個々の分子が実際に互いに浸透する動的構造である。 それゆえ、彼らはお互いに浸透し、互いから離れて横たわっているため、私たちはそれらを見ることができません。 この距離があれば、我々はそれらを見ただろう。 それらは横に並んでおり、個々のヌクレオソームフィブリルを相互に貫通することができ、個々のフィブリルは形成されず、共通の溶融溶液ヌクレオソーム原繊維が存在する。 初代クロマチン原線維はどこから来たのですか?彼らは何十年も前から見てきましたか? ここではすべてが非常に簡単でした。正常なアルデヒド固定を保持し、それを凍った状態で学ぶと、30ナノメートルのクロマチン原線維が見えます。 これは固定の人工物です。 クランプされたフィブリルを固定すると、それが目に見えるという事実のために。 それがカールしていなければ、DNAヒストンタンパク質を使ってコンパクト化された1分子の溶融分子が見えていたでしょう。

 カート
カート