カート
カート

ビタミンD
01 Nov 2016
ビタミンD以前のカルシウム交換調節に非常に受動部品に割り当てられています。 このビタミンは小腸と効果的な行動Parathyroidin(パラトルモン、PTG)の中のカルシウムの吸収を提供するために十分なだけの血で存在しなければならないと考えられました。 今では、カルシウムの交換におけるビタミンDの役割ははるかに活性であることが知られています。それはビタミンと呼ばれているが、実際には、一緒にPTGで血中カルシウム濃度の主なレギュレーターの機能を実行するホルモンです。 実際に、ビタミンDは、ホルモンの典型的な兆候を持っている:1)それは皮膚で合成され、この合成の理想的な条件でそれが栄養との進入が義務であるとすることはできませんです(非常に十分であることができる)、2)、彼が持っていたされていますそれは厳密に調節可能な酵素によって活性化される他の臓器への血液と、3)その活性型(カルシトリオール)は、血液中のカルシウム濃度の上昇に最終的につながる、標的組織の特定の受容体に結合しています。 また、今日では、ビタミンDは、他の機能を実行することが知られています。 カルシトリオール受容体は、骨髄細胞、リンパ球、上皮細胞、膵臓、筋肉や神経細胞の島々の細胞を含む多くの細胞上ご利用いただけます。 これらの受容体の活性化は、カルシウム交換に結合していない様々な効果が続きます。 履歴情報。 ビタミンDのコール2 congenerous脂溶性債 - kholekaltsiferolとエルゴカルシフェロール、防止またはくる病の治癒する能力を有します。 このビタミンの発見前に適度な気候帯の都市に住むくる病多くの子供たちに苦しみました。 一部の研究者は、他の人が配達の特徴で、その理由を見て、くる病が原因で新鮮な空気と日光の不足の発達と考えました。 フェアは、両方の病気が(タラ肝油から)肝油の食事に添加することによって予防またはように硬化されるように管理されるビューのポイント、日光の影響(; Huldschinsky、1919メランビー、1919)でした。 (; Steenbockと黒、1924ヘスとウェインストック、1924年)、1924年には、動物でくる病が紫外線放射だけでなく、個人だけでなく、それらによって消費される栄養を回復することが示されました。 これらの観察は、holekaltsiferolとエルゴカルシフェロールの構造の解釈につながりました。 これは、これらの物質は唯一の生化学的な一連の変換後の活性を得ることも設立されました。 ビタミンDの活性化のプロセスは、一般的にアメリカ(デルーカとSchnoes、1976)と研究者による英語(Kodicek、1974)を検討しています。
化学的性質と情報源
動物と植物由来のいくつかのステロールの紫外線放射は、S-9及びS-10の原子との間の通信の破裂はしかし、同時にすべてではない(必要に応じて、この目的のためであるビタミンDの活性を有する債券に変換します)抗くる病の活動を得るsterola。 動物に見られる主なプロビタミンは、皮膚で合成された7-degidrokholesterinです。 紫外線放射7-degidrokholesterinの影響下kholekaltsiferolになります(D3ビタミン)
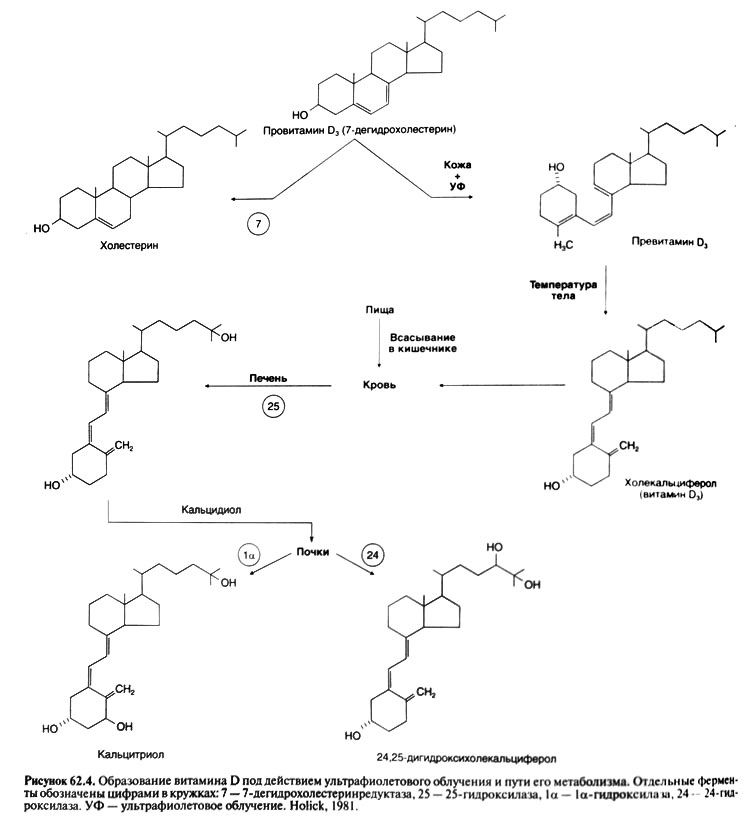
紫外線放射および彼metapainの方法の影響下でのビタミンDの形成。
中間光分解物は、紫外線放射の皮膚に集まるプロビタミンD(D3ビタミン6.1 - シス異性体)が発見された(無なめる、1981)です。 この異性体はゆっくりと自然にD3ビタミンになり、いつか照射後にそのソースにすることができます。 植物のエルゴステロールでの出席者は、D(D2ビタミンの前駆体、またはエルゴカルシフェロール)のプロビタミンを表します。 エルゴステロールおよびD2のビタミンは、S-24原子においてS-22およびS-23原子とmetilnyグループ間の二重通信の有無に7-degidrokholesterinとD3ビタミンからそれぞれ異なります。 ビタミンDは多くのビタミン薬の一部であり、照射されたパンと牛乳で存在します。 以前のビタミンDによって呼び出された物質は、(それは動物ではくる病を防ぐために、ビタミンDとD3のいくつかの種類の能力が強く異なります。くる病の発展を妨害する債券の混合物であったが、一人でそれらの両方がほぼ均等に作用する。また、両方のビタミン私たちは、一般的な用語のビタミンDを指定します
活性化
栄養に到着するビタミンDのように、とだけ一連の変換後の皮膚の利益の生物学的活性で合成。 主活性代謝物 - カルシトリオールは、 - 詳細はホルストとラインハルト(1997)のレビューで、例えば、参照のためにビタミンDの二つの連続ヒドロキシル化の結果として形成されています。 取ることを忘れてはいけないビタミンB12を 、より良い結果を得るため。
ビタミンDのヒドロキシル化
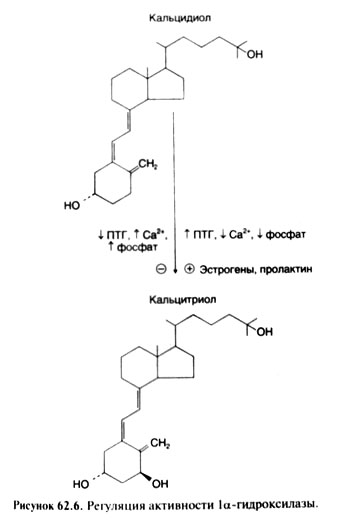
それはcaltsidiolが形成され、その結果としてのビタミンDの活性化の第一段階です。 これは、肝臓で進行します。 反応はNADFN及び分子酸素の存在下でmikrosomalnyまたはミトコンドリア25ヒドロキシラーゼによって触媒されます。 caltsidiolの水酸化。 D - - svyazy-vayushchimタンパク質(およびグロブリン)プラズマが肝臓caltsidiolからビタミンについての接続血液に来ます。 カルシトリオールへの変換 - - ビタミンDの活性化の最終段階は、マクロファージを含む、そのような能力は、他の細胞は、(ライヒェルら、1989)を有しても腎臓で主に発生します。 kaltsidiolのヒドロキシル化は、近位細管の細胞のmitochondrionsに局在1A-ヒドロキシラーゼによって行われます。 NADFNと酸素分子が反応に関与します。P450シトクロム、黄色酵素とferredoksinは、図1a-ヒドロキシラーゼの一部です。 それは、混合機能を持つオキシダーゼを表します。
カルシトリオールの分泌レベルは、血液中のカルシウムの正常な濃度を維持するために十分であったように、図1a水酸化酵素の活性が調節されています。 ビタミンD、カルシウムおよび酵素増加のNatrii phosphas活動の低下、消費では、消費の増加で - 減少します。 また、ラ・ヒドロキシラーゼの活性は、PTGの影響下に拡大され、また、おそらくProlactinumおよびエストロゲン(図62.6)。 ラヒドロキシラーゼの活性の短期および長期の調節の両方が可能です。 その合成変化の後者率です。 PTGの作用は明らかに、一連の段階を通してラヒドロキシ(シーゲルら、1986)のferredoksinovy成分に影響を及ぼすfosfoproteidfosfatazaを活性化する、tsAMFによって媒介されます。 いくつかのデータによると、低カルシウム血症は、PTGの分泌の刺激を介してのみならず、図1a-ヒドロキシラーゼを活性化するだけでなく、即時です。 低リン血症は、図1a-ヒドロキシラーゼの活性の鋭い立ち上がり(;フレイザー、1980;ローゼンとチェズニー、1983 Hausslerとマケイン、1977)につながります。 負のフィードバックのメカニズムにカルシトリオールは、それが腎臓に、およびPTGの形成の抑制を通じて即時であるとして活躍し、この酵素の活性を減少させます。 図1a水酸化酵素の活性に対するエストロゲンとProlactinumの影響のメカニズムは知られていません。
生理学や薬理学
ビタミンDの主な効果は、血液中のカルシウム濃度の上昇しています。 ビタミンDの影響下Natriiのphosphasの交換はカルシウム交換と平行に変更されます。 同時に、より多くのデータは、ビタミンDの役割を示し、多くの他のプロセスに(下記参照します)。
1)、小腸でこれらのイオンの吸収が激化骨から2)それらの放出、それらの腎排せつの3)うつ病:ビタミンDは、以下のメカニズムによって、血漿中のカルシウムとNatriiのphosphasの正常な濃度を維持しています。 骨組織のmineralizationsにおけるビタミンDの直接の役割を推定することは困難です。 それは血漿中、カルシウムと骨形成のために十分なNatriiのphosphasの濃度のみを提供すると考えられました。 しかし今では、ビタミンDは、骨組織の更新に関与する細胞の両方の間接的な、および直接的な影響をレンダリングすることが知られています。
カルシトリオールの作用のメカニズムは、ステロイドおよび甲状腺ホルモンと同様です。 カルシトリオールは、細胞標的の細胞内受容体に結合し、そしてホルモンである - 受容体複合体は、遺伝子の転写を変化させる、DNAと相互作用します。 Kaltsitriolovy受容体は、ステロイドおよび甲状腺ホルモンの受容体(;パイク、1992エヴァンス、1988)のように、スーパーファミリーにほかに属しています。 カルシトリオールはまたbystry(どうやら、余分なゲノム)の効果(Barsonyandマルクス、1988)を引き起こします。
腸内のカルシウム吸収 。 ビタミンDの欠乏ラットにおける腸内でのカルシウムの吸収の乱れが50年以上前に明らかにされています。 4時間漿液カバーに粘液、腸管からのカルシウムの動きを加速 - 2はすでにビタミンの活性形のような動物による紹介。 この効果のメカニズムは(ワッサーマン、1997)絶対に明確ではありません。 家族小さなカルシウム結合タンパク質(カルビンジン)から1タンパク質の比較的誘導が早期に観察されます。 一部の研究者は、腸細胞の基底膜への刷子縁を通ってカルシウムとその拡散の移転を促進するものカルビンジンを検討してください。 その他には、カルシウム輸送(Nemereとノーマン、1986,1988)へのカルビンジンのレベルの不調和を指摘し、そのカルシトリオールは、腸の内腔からのカルシウムが腸の細胞の小胞に到達し、これにより、エンドサイトーシスを刺激すると信じています。 これらの小胞は、それはまた、血液に入るところから側底膜にカルシウムを提供リソソームと合併(Cancelaら。、1988)。 カルシトリオールはカルシウムのような輸送を強化する手段により、メカニズムは知られていません。 血液中の腸細胞からのカルシウムの放出は(ワッサーマン、1997)拡大されるカルシトリオールの影響下での数の細胞膜のカルシウムポンプによって行われます。 Dのビタミン欠乏症を有する動物におけるカルシトリオールの効果の開発の面では、膜受容体を介して、おそらく働いて、しかし、ビタミン欠乏症のカルシトリオールの欠如のために、その作用のゲノム機構に証言し、輸送の立ち上がり(数分以内)に高速せることも可能ですカルシウム(Cancelaら、1988)。 骨からのカルシウムの動員。 骨のミネラル物質のビタミン欠乏Dの含有量を有する動物で明らかに減少しているが、直接石灰化を促進するために、このビタミンの能力が証明されていないままです。 ビタミンDが原因腸(幹、1980)、これらの物質の吸収の増強にのみ、血液中のカルシウムとNatriiのphosphasの濃度を維持することによって骨の正常な石灰化を提供すると考えられます。 本当に、ビタミン - 子供でIIのようなD-依存くる病が正常にカルシウムとNatriiのphosphas(下記参照)の投与により静脈扱われます。 逆に、ビタミンDの生理学的用量は、離れて、骨からのカルシウムの洗浄を強化し、高用量では大幅に骨組織の更新を促進します。 骨組織の再吸収にカルシトリオールのリモート副甲状腺の影響力を持つ動物で弱体化しているが、高リン血症の補正は、この影響(茎、1980)を復元します。 したがって、PTGおよびカルシトリオールは、互いに独立して、骨組織の吸収を強化します。
骨組織の更新にcaltsitriolの刺激影響のメカニズムは部分的にしか研究されています。 多くの相互接続された要因は、(Hausslerは、この影響で1986に参加。ライヒェルら、1989)ことが確立されています。 成熟破骨細胞は、明らかに、しかしcaltsitriolは、成熟osteoklastの機能のこれらのケージで再吸収センターにケージの前任者の動きだけでなく、買収を刺激し、caltsium受容体を奪われcaltsitriolに敏感ではないされている(Mimuraet、1994)。 caltsitriolに、また再吸収を刺激する他の物質に対するこの反応のシャープ違反は(誰に骨組織の再吸収が遅くなる)大理石骨病の特徴です。 、およびシルト-1(tsitokin - カルシウム受容体は骨芽細胞で利用可能であり、caltsitriolはosteocaltsin含むタンパク質の数、これらのケージ(酸carboksiglu-taminで遺跡を含むmin.ツー依存性タンパク質のVit)で開発を刺激します骨組織の再吸収)(スピアら、1988)を強化します。
腎カルシウムの排せつとNatrii phosphas
ビタミンDは独立して、カルシウムとNatrii phosphasの排せつを低減します。 おそらく、それが近位細管での再吸収を強化します。 ビタミンDのこの効果の値は不明瞭なままです。
その他の効果
今では、カルシウム交換カルシトリオールの影響だけではないことが確立されています。 Calcitron受容体は、多くの細胞(パイク、1992)で存在しています。 サイトカインの成熟カルシトリオールの影響下およびリンパ球と単球の分化が加速し、また、生産は(免疫系へのカルシトリオールの影響に関する詳細はAmento、1987年のレビューで参照)が発生します。 特に関心の増殖を抑圧するために、腫瘍細胞(バンLeeuvenとPOLS、1997)の分化を引き起こすためにカルシトリオールの能力によって吸引されます。 細胞にこのアクションを維持し、血漿中のカルシウム濃度の上昇を引き起こすことはありませんが、カルシトリオールの類似体の検索が開始されます。 このような物質は、抗腫瘍剤として使用することができます。 カルシトリオールはkeratinotsitの増殖を圧迫し、それらの分化を加速するように、おそらく、それは乾癬(Kragballe、1997)の治療に応用されます。
老人性認知症とアルツハイマー病の発症のダブルス危険性よりも、より高度な年齢で、ビタミンDの欠乏、仕事神経学雑誌で公開されて見つけた研究者の国際的なグループ。
血液中のビタミンDの含有量及び脂肪組織の量との間の依存性が証明されています。 より多くのあなたのビタミンDは、脂肪分が少ないとより多くの筋肉のバルクを持っています。実験室での研究は、筋細胞はまた、それは、ビタミンDの欠乏と筋肉細胞が迅速に筋肉組織を分割することが明らかになったビタミンDの影響で少なくMyostatinumを作ることを示しました。
カルシトリオールへの抗エストロゲン効果を検討したその間 - スタンフォード大学の科学者たちは、ビタミンDに専念動物の研究を行いました。 Endokrinologiya誌に発表された結果は、カルシトリオールの影響一般的に脂肪組織がボディビルダーに興味深いものになることができると言います。 それに、これは����������、レトロゾールおよび����������などの抗エストロゲンの効果を強化します。 ヒト細胞においてOestradiolumの生産は生物においてカルシトリオールの濃度の増加の過程で減少することが判明した人々に、マウスの研究の結果を転送することが可能である場合。 この事実はOestradiolumの小さい量は、脂肪細胞の身長の遅延を意味するボディビルダーに興味深いもの、また、それらのサイズを小さくすることができます。 また、女性化乳房の出現のうつ病。 実験は、ビタミンDの受信arimideks、ekzemestanとレトロゾールの効果を強化することが示されました。 西洋文明の代表は、原則として、血液中のビタミンDの濃度が低すぎるを持っているとして、このビタミンを含有添加剤の受信とそれを組み合わせる場合はAASを使用して、選手がアロマターゼ阻害剤の追加の効果を得ることができると仮定することが可能です。
吸収、代謝および排泄
ビタミンDは、通常、内部に規定され、そしてほとんどの場合、十分にまで浸漬します。 D2のビタミン、およびD3ビタミンの両方が小腸に吸い上げているが、最後のわかりやすさは、それは、上記の可能性があります。 ビタミンDの最大数がアップ浸される小腸のどの部門では、自然食品によって異なります。 浸しアップビタミンDの主要部分は、初めにリンパ液中のカイロミクロンの一部として発見されました。
ビタミンDの吸収の過程で重要な役割は、それdesoxycholeic酸で特に存在する胆汁で再生されます。 したがって、肝臓の病変またはコールの方法でビタミンDの吸収が著しく破壊されます。
ビタミン - - について - 結合タンパク質浸しアップビタミンDはarglobulinyプラズマに接続します。 25時間が、脂肪組織における - - プラズマへのビタミンB 19を作る非常に長いです。
- - 結合タンパク質、それは知覚になっ既にとしては、肝臓のビタミンDにもビタミンについての接続caltsidiolになります。 Caltsidiolは、初期接着よりも、このタンパク質に対してより大きな親和性を有しています。 Caltsidiolは、血液中のビタミンDの主な形態です。 そのT1 / 2から19日。 骨組織の更新のPTGレベルと加速度の立ち上がりが25 ng / mlでより低い濃度で既に生じ得るもののng / mlで50 - caltsidiol者濃度で15に対して垂直なピア・プラズマです。 カルシトリオールT1 / 2は、3〜5日に人のピアプラズマです。 入力された用量の40%が10日間に除去される(Mawerら、1976)。 24,25-digidroksikholekaltsiferolに - 腎24ヒドロキシラーゼは、カルシトリオール(1,25-digidroksikhapekaltsiferol)1,24,25-trigidrok-sikholekaltsiferolで、かつcaltsidiol(25-gidroksikholeka-ltsiferol)をオンにします。 この酵素は、カルシトリオールによって誘導され、図1A-ヒドロキシラーゼを刺激因子によって阻害されます。 1,24,25-trigidroksikholekaltsiferol、および24,25-ジgidroksikholekaltsiferolとしてはカルシトリオールよりも、小さい活性を有し、そしておそらく排泄代謝物を表します。 カルシトリオールの代謝は、側鎖の酸化によっても行われます。
ビタミンDは、入力されたビタミンの小さな部分が発見された尿に、胆汁と一般に除去されます。 ビタミンD及びその代謝物は、従って、小腸の炎症を患っているか、その大規模なサイトの切除を転送され、腸の吻合を伴う患者でアクティブ腸肝回路にさらされている、ビタミンの吸収の乱れは、ビタミン欠乏につながります。
PhenytoinumとFenobarbitalumとビタミンDの相互作用の重大な結果を説明します。 それは長い抗けいれん薬を受けている患者では、くる病とosteomalacyは時々観察されました。しかし、これらの薬剤は、骨組織の迅速な更新と骨粗鬆症の開発を促進し、腸でのカルシウム吸収を低下させる、より多い(ワインスタインら、1984)。 このような薬剤を受け入れた患者では、血漿中のcaltsidiolの濃度は、したがって、それはPhenytoinumとFenobarbitalumが不活性代謝物(ハーンら、1972)にビタミンDの転換を加速することを想定した削減されます。 しかし、抗痙攣薬を受けた患者の大部分では、血漿中のカルシトリオールの濃度が正常なままである(Jubizら、1977)。 これらの薬剤はまた、ビタミンKの肝代謝を促進し、例えば、osteokaltsinをビタミンに依存性タンパク質の合成を遅くします。
要件および測定の単位
ビタミンDの予防ニーズに関するデータの徹底的な報告書は(1963年栄養に関する委員会)米国小児科学会の配信に委員会によって行われました。 Mellanbiが(タラ肝油から)タラ肝油によるくる病の予防の可能性を確立した1919年以来、長年そこに渡されました。 今、この病気はめったにアメリカで満たしていません。 ビタミンDの追加の受信における必要性があるなぜ熱帯諸国における場合くる病の予防は、皮膚の日射量は軽微であり、特に冬には、温暖な気候を持つ国では、太陽放射によって提供されています
以前の特別な栄養補助食品は、したがって、どの年齢でも、その過剰消費の危険がある今、このビタミン食品(特に乳製品、製品細断穀物やお菓子から)で豊かにビタミンDの毎日のニーズの満足のために必要でした。 ビタミンDの追加の受信のためだけでなく、年齢や(例えば、妊娠またはlactemia用)生物の状態に依存するだけでなく、栄養の構造にする必要があります。 ビタミンの過剰摂取は重大な結果につながることができます。 子供でのみ1800 MEのビタミンD(下記参照)は、本体の高さを遅くすることができます。 したがって、それだけダイエットの勉強を慎重後にビタミンの追加の受信をお勧めすることが必要です。
関係なくビタミンDは400 ME /日の用量での消費が完全にくる病と(早産を含む)新生児の正常な身長の予防を提供して供給します。 この量は、ティーンエイジャーを含むあらゆる年齢層の人々のために、おそらく十分です。 一部のデータは、妊娠とlactemiaでビタミンDの必要性の上昇を示しますが、400 ME /日は、これらの状態でも十分でなければなりません。
国際単位は(ME)holekaltsiferolの0025 MKGの生物学的活性と同等である(つまり、holekal-tsiferolの1 mgで40 000 MEをサポートしています)。
ビタミンD及びその誘導体の以前の生物学的活性は、くる病を防止する能力により決定しました。 このような方法はまだ実験的な目的で使用されています。 ビタミンD誘導体。 かなりの関心が実験や診療所でのビタミンDの一連の誘導体の使用の可能性に惹かれています。 そのうちの一つ - Dihydrotachysterolumは - 復元されたD2のビタミンを表します。 その構造式は、以下の、
抗くる病活動にDihydrotachysterolumは約450倍弱いビタミンDよりもですが、Dihydrotachysterolumがで血液中のカルシウムの正常な濃度の維持に適用される。このような理由からビタミンDよりも、はるかに強い高用量で骨からのカルシウムの動員を刺激しますhypoparathyrosis。
Dihydrotachysterolumは、明らかに教育25 hydroxydihydrotachysterolumsとヒドロキシル化され、そして腸と骨に影響を与えます。 この物質は、作品やリモート腎臓を有するラットで - 手段、その活性は、腎臓で1-&